Méthode de Turing et morphogénèse : illustration des équations de réaction-diffusion
Auteurs:
El Mellah Ileyk, Loh Alan, Pottier Alizée
Date de création :
2012
Date de mise à jour :
13 décembre 2012
Table des matières
Liste des paramètres d'entrée de l'applet
-
titre : Machine
label : machine
Choix de la puissance de votre machine, ce qui détermine la taille des grilles et donc a une incidence sur le temps de calcul.
Note: pour la machine puissante, éviter de solliciter l'affichage des quatre panneaux à la fois.
Valeurs possibles :
Modeste Moyenne Puissante
-
titre : Rayon act
label : r1
Valeur du rayon de diffusion de l'activateur, R1
-
titre : Rayon inh
label : r2
Valeur du rayon de diffusion de l'inhibiteur, R2
-
titre : Diff act
label : diff1
Valeur du coefficient de diffusion de l'activateur, J1
-
titre : Diff inh
label : diff2
Valeur du coefficient de diffusion de l'inhibiteur, J2
-
titre : Offset
label : offset
Valeur initiale du palier (équivalent au rapport des concentrations à la source), h
-
titre : Grille
label : grille
Choix de la grille d'évolution des taches. Attention à ne pas utiliser de maillage de VORONOY avec la géométrie queue!
Valeurs possibles :
Simple Hexagonale Voronoy
-
titre : Géometrie
label : geometrie
Définition des conditions au bord (ie de la géométrie globale, carrée ou rectangulaire). Attention à ne pas utiliser de maillage de VORONOY avec la géométrie queue!
Valeurs possibles :
corps queue
-
titre : Durée
label : nstep
Durée de la croissance
-
titre : synchro
label : synchro
Evolution synchrone, asynchrone random ou semi asynchrone
Valeurs possibles :
semi-asynchrone asynchrone synchrone
-
titre : powerlog
label : powerlog
Puissance du spectre logarithmique (pour voir les harmoniques) ou non (pour un meilleur contraste)
Valeurs possibles :
linear spectrum logarithmic
-
titre : Couleur active
label : couleur1
Couleur des cellules activées (doit être différente de la suivante!)
Valeurs possibles :
rouge orange jaune noir blanc bleu vert
-
titre : Couleur passive
label : couleur2
Couleur des cellules non activées (doit être différente de la précédente!)
Valeurs possibles :
rouge orange jaune noir blanc bleu vert
-
titre : Fraction Voronoy
label : pointsvoronoy
Rapport du nombre de cellules de DELAUNAY par le nombre de pixels à l'écran (ie, en géométrie corps, dimx^2)
Valeurs possibles :
0.01 0.07 0.2 0.5
Liste des paramètres de sortie de l'applet
-
label : taches
-
label : lc
-
label : lcnow
-
label : lcderiv
-
titre : lc_x
label : lcx
-
label : fft
-
label : somme
-
label : tpsactuel
-
label : texteapplet
-
label : lcfourier
-
titre : pulsation spatiale
label : pulsationspatiale
-
titre : spectre
label : spectre
Introduction
On remarque dans la nature l'émergence de structures complexes à plusieurs échelles qu'on a peine à croire explicables par des règles simples. Les taches formées sur les pelages d'animaux comme le guépard, ou le lynx, ou encore sur la structure des couleurs des plumes de certains oiseaux ou les carapaces d'insectes en sont quelques exemples.
C'est en s'intéressant à cette problématique qu'Alan Turing, mathématicien et informaticien britannique publia en 1952 l'article
The Chemical Basis of Morphogenesis (Les Fondements chimiques de la morphogénèse). Dans cet article est proposée une théorie d'émergence de structures plus ou moins complexes à partir d'un modèle de réaction-diffusion.
Guépard dans la brousse

Crédit : KouK's (Flickr, licence Creative Commons)
Sur cette page est décrite l'application de ce modèle dans le cadre d'une appliquette Java (qu'on peut trouver
ici).
Champ
L'appliquette sert surtout dans le cadre de la biologie théorique, voire la géographie ou la géologie si l'on s'intéresse à l'étude de la formation des dunes.
Il n'entre donc pas vraiment dans le cadre astrophysique, du moins en l'état, à deux dimensions.
-
Objectifs
-
L'objectif sera de mettre en évidence l'émergence de structures complexes à partir de règles simples. L'accent a été mis sur l'analyse de grandeurs caractéristiques
des structures obtenues, et des outils ont été développés dans ce but.
Rappels de cours
Comme le modèle étudié vise à reproduire la pigmentation dans le règne animal et à expliquer les structures variées qui y règnent, il semble nécessaire de rappeler quelques notions à ce sujet.
Ces rappels n'en seront vraiment que pour ceux qui ont déjà suivi quelques cours de biologie. Il est possible qu'il s'agisse de nouvelles notions pour les lecteurs physiciens.
La couleur noire a souvent été choisie comme couleur de cellule activée dans l'appliquette. La mélanine est en effet dans le règne animal l'un des principaux colorants des téguments (qui font le lien entre l'intérieur et l'extérieur d'un organisme vivant).
Il n'y a pas de formule chimique pour la mélanine car celle-ci est un agrégat de plusieurs types de macromolécules. A titre d'exemple, voici la formule de l'une
d'entre elles, l'eumélanine.
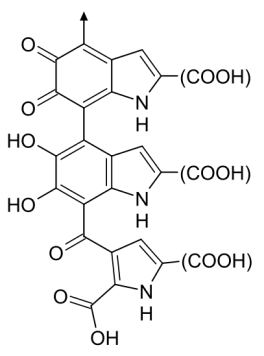
Roland Mattern
Les différents types de mélanine peuvent être à l'origine de nombreux tons de couleur.
Outre cet exemple, il existe de nombreux autres pigments à l'origine des couleurs des organismes vivants. L'idée est que toute cellule productrice du pigment
n'est pas activée ou désactivée de façon permanente, mais le produit en fonction de stimuli qu'elle reçoit de son environnement sous forme d'enzymes.
Ainsi la tyrosinase est une enzyme activatrice responsable de la production de mélanine.
Bibliographie
Wikipedia, page sur la mélanine et la tyrosinase.
Lien sur la tyrosine.
Explications
Description du Modèle
Le modèle de Turing repose sur l'interaction entre éléments infinitésimaux, les cellules. Celles-ci peuvent être considérées comme analogues aux cellules vivantes de la peau d'un animal ou d'un embryon d'animal, par exemple. Dans un autre domaine, on pourrait considérer des grains de sable s'organisant dans des dunes.
On considère un réseau, une grille, de cellules qui peuvent être dans deux états possibles: un état activé ou désactivé. Cela s'accorde bien avec le fait que chez les mammifèes par exemple une tache noire serait dûe à l'activation de mélanine qui colore la peau.
On part de l'idée de la coexistence de deux types de sécrétions pour les cellules :
- Un effet activateur à courte portée
- Un effet inhibiteur à longue portée
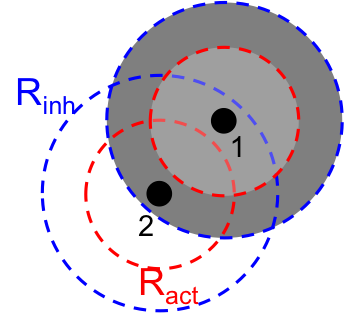
Ces deux effets, issus des cellules elle-mêmes, font que l'état d'une cellule à un instant donné dépend de celui des cellules autour d'elle.
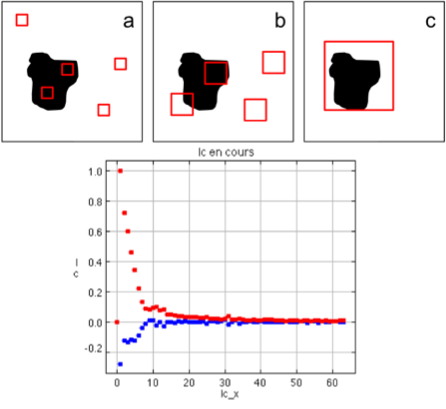
On peut le voir sur la figure ci-dessous: la cellule 2 reçoit l'inhibiteur de la cellule 1 (car elle est dans sa zone gris foncé, sa distance à la cellule 1 est donc inférieure à R2).
Cependant la cellule 2 est hors de la zone d'activateur sécrété par la cellule 1 (zone gris clair), car sa distance à la cellule 1 est supérieure à R1. Elle n'est donc pas incitée à s'activer par la cellule 1.
Rayons d'action
Equation d'Evolution
Ces équations traduisent la diffusion de l'activateur et de l'inhibiteur sécrétés par une cellule et affectant ses différentes voisines.
On note :
- R1 le rayon d'action de l'activateur
- R2 le rayon de l'inhibiteur
- J1 le coefficient de diffusion de l'activateur
- J2 le coefficient de diffusion de l'inhibiteur
- h le palier d'activation
![cellule_i(t+1) = signe\Biggl[h + J1 \sum \limits_{r_{ij}<R1}{cellule_j(t)} + J2 \sum \limits_{R1 \leq r_{ij}<R2}{cellule_j(t)}\Biggr]](eq_tex_AutomateTuring_website/equation1.png)
La formule ci-dessus signifie qu'on va examiner l'état des cellules (j) dans un rayon R1 pour prendre en comptre leur effet activateur et l'état des cellules (j) dans un rayon R2 pour prendre en comptre leur effet inhibiteur. Cela sert à calculer l'état de la cellule (i) qui est la cellule en cours d'actualisation.
Modes de mise à jour de la grille
On en a considéré trois, complémentaires.
Le mode asynchrone actualise les cellules une par une de façon aléatoire, le paramètre "durée" de l'appliquette déterminant le nombre de cellules actualisées avant chaque nouvel affichage.
Le mode synchrone calcule les états de toutes les cellules de l'image (n+1) à partir de celles de l'image n simultanément.
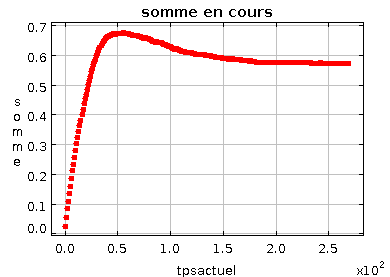
Le mode semi-asynchrone est le plus utile: il actualise l'image en calculant l'évolution cellule par cellule de façon asynchrone, mais considérant aléatoirement les cellules, il n'affiche la prochaine image que lorsque toutes les cellules ont été actualisées de la sorte. Cela permet d'éviter la présence d'une première constante de temps lors de l'apparition des structures qui n'était que le temps nécessaire pour visiter aléatoirement toute la grille.
Ci-dessous, on peut voir la stabilisation d'une image lors de l'apparition des taches; l'ordonnée représente la somme des valeurs de toutes les cellules (la valeur +1 est associée à une cellule activée, -1 à une cellule désactivée). Cette somme permet de suivre la stabilisation de l'automate au niveau global.
Stabilisation
Intérêt des Géométries
Les cellules étudiées sont placées sur des canevas, des maillages les liant entre elles.
Cependant on peut voir ci-dessous un exemple d'émergence de structure de type empreinte digitale avec de forts effets horizontaux et verticaux: l'anisotropie est issue de la structure de la grille simple (rectangulaire). Donc la nécessité d'introduire d'autres structures apparaît.
simple

Structure fine de grille simple (Ract=1,Rinh=2, Jact=1, Jinh=-0.1, Offset=0)
Voici ce que donne la grille hexagonale. On voit que les structures horizontales et verticales liées à la géométrie de la grille simple ont disparu. Les structures sont également plus épaisses, ce qui est encore une preuve de l'influence du rayon d'action de l'inhibiteur sur la structure.
Hexagonale

Premier exemple de grille hexagonale (Ract=1,Rinh=5, Jact=1, Jinh=-0.1, Offset=0)
Un troisième maillage, celui de cellules de Voronoï (ou triangulation de Delaunay), vise à s'affranchir des artefacts dûs à la régularité du maillage.
Voronoy

Structure de maillage non régulier (Ract=3,Rinh=4, Jact=6.5, Jinh=-3, Offset=0)
Pour en savoir plus sur ces différents maillages, voir ici :
Maillages. Ces grilles ont également été utilisées pour développer
une appliquette concernant
le modèle d'Ising.
Outils d'Analyse
Dans le but de déterminer la taille caractéristique des taches d'animaux ainsi obtenues, deux méthodes ont été développées. La première est basée sur l'étude de la fonction de corrélation de la grille et la deuxième sur l'analyse de la transformée de Fourier de l'image obtenue.
Fonction de Corrélation
Le calcul de la fonction de corrélation des taches se base sur une méthode simple consistant à faire grossir un élément structurant et analyser "ce qui passe au travers". Par exemple, on peut déterminer la taille des pores d'une passoire en testant le passage de grains de sable de différents diamètres. La courbe représentative des grains ayant traversé en fonction de leur diamètre est maximale pour des grains de petite taille, puis diminue car des grains se bloquent mutuellement. Elle ne devient nulle que lorsque les grains ont atteint un diamètre tout juste supérieur à celui des pores de la passoire.
Cet effet est celui qui a été exploité ici. Une cellule activée (noire, par exemple) se voit affectée d'une valeur égale à
, tandis qu'une cellule non activée reçoit la valeur
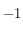
. On fait alors grossir un carré de taille
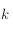
(avec
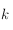
variant de 1 pixel à la moitié de l'image), que l'on déplace à travers toute l'image (il y a
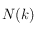
déplacements qui dépendent de
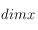
et
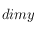
correspondant aux dimensions de la grille étudiée). A chaque déplacement, on additionne les valeurs des cellules incluses dans cet élément structurant (en valeur absolue et en normalisant par le nombre de cellules :
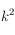
). On reporte la moyenne de toutes les sommes correspondant à tous les déplacements du carré en fonction de sa taille
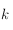
(voir Figure ci-dessous).
Exemple de fonction de corrélation
La courbe rouge représente la moyenne de la somme des cellules pour tous les déplacements d'un carré de taille
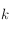
en fonction de
. La courbe bleue est sa dérivée calculée grossièrement comme étant le taux de variation entre les points consécutifs.
On part de petits carrés de taille 1 pixel (le premier point de cette courbe est à 0 mais n'a bien entendu aucune signification car un élément structurant de taille nulle n'est pas défini), cette situation est schématisée en (a). Les cellules incluses dans un carré de 1 pixel de coté sont forcément au nombre d'une unité :
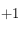
ou
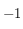
, ce qui, en valeur absolue et une fois normalisé, donne le résultat attendu
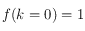
.
Plus la taile du carré croît, et plus la somme de son contenu diminue (situation (b)) : les valeurs des cellules activées et non activées commencent à se compenser mais une composante domine toujours l'autre. Arrive alors le moment où la taille du carré, peu importe sa position sur l'image, englobe quasiment autant de cellules activées que de désactivées (situation (c)). La courbe rouge rencontre alors une inflexion relativement bien prononcée car dès lors, la croissance du carré ne change pas fortement la valeur de la fonction de corrélation.
On remarque alors la présence de deux régimes distincts : une forte décroissance de la fonction de corrélation suivie d'une période plus plate où elle se rapproche d'une valeur constante (qu'on peut noter
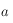
). Ce genre de courbe peut être approximée par une fonction de type
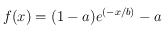
. Cependant, l'ajustement d'une telle fonction sur ces points expérimentaux pour déterminer les coefficients a et b n'est pas une chose aisée. Pour contourner cette difficulté, deux ajustements linéaires ont été préférés. On détermine le point approximatif séparant les deux ajustements en utilisant la dérivée de la fonction de corrélation (courbe bleue) : on définit ce point approximatif
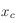
là où elle s'annule.
Ensuite, on réalise un ajustement linéaire par la méthode des moindes carrés sur les points compris entre
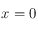
et
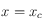
. Sur la deuxième série de points, une moyenne de leurs valeurs est calculée et donne directement l'ordonnée à l'origine d'une fonction affine constante. L'intersection entre ces deux droites détermine la taille caractéristique du système issue de la fonction de corrélation.
Transformée de Fourier

Transformée de Fourier (TF) en échelle logarithmique de "Réseaux imbriqués" avec une maille hexagonale
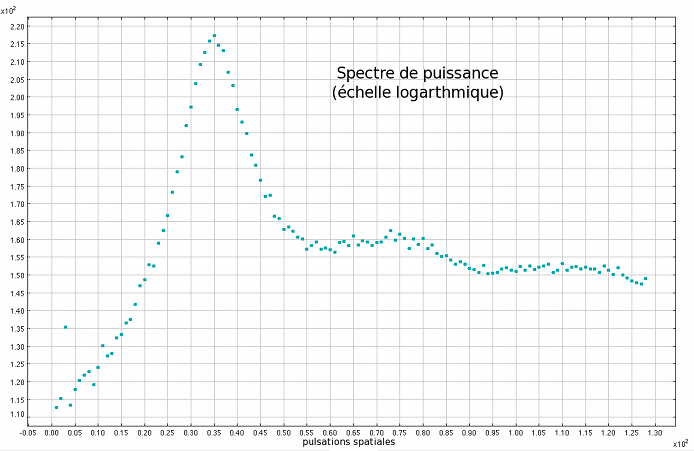
Section moyennée sur tous les angles de la TF précédente
Pareille aux figures d'interférences lumineuses qui se construisent peu ou prou par coïncidence d'ondes électromagnétiques modulées par un masque, la figure ci-dessus, appelée transformée de FOURIER (ou TF), est une représentation dans l'espace de FOURIER du motif qui se développe. Les zones lumineuses et sombres correspondent à des interférences constructives et destructives rendues possibles grâce aux périodicités de la figure transformée. Ainsi, la transformée de FOURIER nous renseigne sur les façons dont se répètent des motifs caractéristiques qui, bien souvent, échappent à la vue.
Hormis les directions privilégiées introduites par le choix du maillage (cf dernier paragraphe de cette section), on n'observera d'anisotropie claire dans le motif que pour certaines configurations bien spécifiques. Cela étant, on a réalisé un moyennage anneau par anneau de la courbe d'intensité de la TF afin de pouvoir la représenter en fonction de la distance au pixel central*1 (cf le spectre de puissance ci-dessus). Si l'interprétation de la TF nécessite une certaine capacité de lecture dans l'espace de FOURIER, ce genre de section intégrée sur les angles permet d'avoir un aperçu plus rapide des périodicités du motif. Ce spectre de puissance (en échelle logarithmique pour mieux faire ressortir les rebonds successifs appelés harmoniques au-delà du 1er pic - le fondamental) est le résultat d'une telle opération pratiquée sur une TF analogue à celle ci-dessus. On voit clairement apparaître le fondamental (premier anneau éclairé sur la TF) qui peut être précisément localisé dans l'espace de FOURIER, en termes de pulsations spatiales. Ces dernières sont reliées aux dimensions dans l'espace direct (en pixels) par la relation suivante :
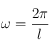
où l correspond à la dimension spatiale associée à cette pulsation. Cette relation indique que plus les dimensions évoquées dans l'espace direct sont grandes, plus les pulsations spatiales correspondantes sont petites. De cette façon, les à-coups et autres discontinuités dans le motif contribueront à augmenter les valeurs de la TF à hautes pulsations alors que les grandes structures contribuent à la TF aux basses pulsations. D'après la définition ci-dessus, L (la taille totale de l'échantillon dont on étudie la TF) fixe donc la résolution de la TF (impossible de voir d'éventuels pics de moins d'un pixel de large sur cette figure!). Ceci s'explique naturellement par l'impossibilité d'observer, dans l'espace direct, la répétition d'un motif plus grand que la zone étudiée*2. Ainsi, cette figure nous permet d'affirmer qu'il existe une fréquence spatiale caractéristique, donnée par le maximum de la figure ie le 1er pic, qui rend compte de la répétition d'un motif bien spécifique de dimension ~ 35 pixels (l'abscisse du fondamental). Comme pour une fonction sinus, il arrive qu'une fonction périodique ait une allure similaire sur 2 demi-périodes successives*3 (ou plus généralement, sur n n-ièmes de période) ce qui "leurre" la TF et crée des maximas locaux dans la figure ci-dessus (au nombre de 2 ici). Ces harmoniques nous renseignent sur la régularité du motif, mais celui-ci peut lui-même comporter des périodicités internes! Bien que la résolution de cette applet aura tôt fait d'entraver la route de celui qui mettra sérieusement cette proposition à l'épreuve, les quelques points qui ponctuent les grandes structures du hanneton foulon ne sont sûrement pas étrangers aux soubresauts que connaît la section aux hautes pulsations.


Deux TF du même motif sans périodicité, mais avec un maillage différent
Les 2 figures juste ci-dessus illustrent l'influence de la maille sur la TF (et donc aussi, bien que de façon moins évidente, dans le motif en lui-même!). Aux grandes pulsations, on observe le comportement du motif aux échelles les plus petites fixées ici par le pixel dont la géométrie définie un maillage. Naturellement, la forme du maillage se fait sentir ainsi que les pulsations qui lui sont inhérentes. Ici, les périodicités ne sont ni spatiales ni temporelles mais angulaires ; en effet, un carré reste inchangé par des rotations de
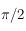
(=> 2 directions privilégiées) et un hexagone reste inchangé par des rotations de
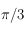
(=> 3 directions privilégiées). Ces propriétés se retrouvent dans la TF où on reconnaît les axes de symétrie de la maille (on laissera au lecteur le loisir de deviner quelle TF est issu de quel maillage…). Parce qu'il comporte plus d'axes de symétrie, le maillage hexagonal rend plus compte d'une modélisation isotrope de la morphogénèse. A haute pulsation, il convient alors de lire les résultats avec précaution afin de faire la part des choses entre les périodicités du signal auquel on s'intéresse et celles de la stratégie d'échantillonnage choisie par l'observateur. Pour reprendre l'analogie exoplanétologique, avec un satellite comme Kepler qui intègre la luminosité sur près d'une demi-heure, malheur à celui qui s'intéresserait à des corps sur une orbite de quelques heures sans prendre garde aux problèmes d'intégration… En revanche, dès que le motif comporte des périodicités de taille bien supérieures à la cellule élémentaire, on observe que, dans la TF, les artefacts dues aux symétries de la maille sont noyés dans le signal issu de ces périodicités (cf la première des deux TF de la configuration guépard des savanes). Pour ce qui est du cas intermédiaire, on peut consulter la deuxième TF ci-après où se mêlent TF des périodicités du motif et de la maille (dans le cas général, on parle de "convolution" entre le maillage, le signal et la fenêtre de mesure).


Deux TF de la configuration guépard des savanes, en maille carrée et hexagonale
N.B. : le premier "F" de "FFT" (pour Fast FOURIER Transform) explique pourquoi les grilles sont carrées (si l'option corps est choisie en entrée, autrement la TF n'est pas calculée) et de taille une puissance de 2. En effet, comme esquissé plus haut, la transformation de FOURIER est une opération qui jouit de propriétés de symétrie très riches et qu'il serait dommage de ne pas exploiter. En "pliant successivement en 2 la TF" (un nombre de fois d'autant plus grand que la taille de la grille est proche d'une puissance de 2), on s'évite des calculs numériquement chronophages en devinant des coefficients par symétrie.
*1 pour les puristes, la TF ci-contre est une translation de la moitié de la dimension du motif dans les deux directions et n'a pas été normalisée (mais la moyenne du motif a été soustraite avant de réaliser la transformation de FOURIER).
*2 de même, l'exoplanétologie suggère une analogie temporelle éloquente : si des satellites comme Kepler ou Corot observent en continu des étoiles en espérant déceler une diminution de leur luminosité due au transit d'une exoplanète, c'est pour disposer d'une base de temps (analogue temporelle de L ci-dessus) suffisante pour détecter la répétition d'un transit d'orbite en orbite (dont la durée - 1 an dans le cas de la Terre - est l'analogue temporelle de la période spatiale de 35 pixels ci-dessus).
*3 on notera que
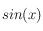
et
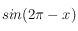
ne diffèrent que d'un moins…
Quelques résultats
Robustesse
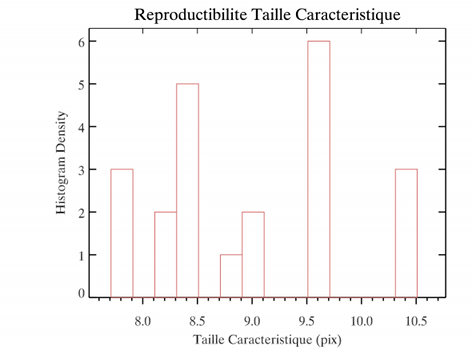
Pour tester la reproductibilité des mesures, on peut relancer plusieurs fois la simulation pour différents paramètres. Comme le montre la figure ci-dessus,
qui représente la distribution de longueurs de corrélation pour plusieurs itérations pour les mêmes paramètres, il existe un effet de discrétisation. Cependant
celui-ci peut être plus dû aux effets de calcul de la longueur par ajustement sur la courbe qu'à un effet d'états initiaux aléatoires.
Taille des taches
Comme il se doit, l'applet Java permet de modéliser des taches semblables à celles d'un guépard (mais pas celles d'un léopard, plus complexes!). En témoigne le set Guépard des savanes qui donne, après quelques itérations, un pelage tacheté. Les longueurs de corrélation et de périodicité calculées s'accordent autour d'une dimension caractéristique d'environ 7 pixels pour cette configuration mais il est possible de modifier cette longueur. En considérant les rayons de l'activateur et l'inhibiteur, nous avons esquissé une étude de l'espace des paramètres (voir section suivante). Par exemple, le set de paramètres ci-dessous donne des taches de dimension beaucoup plus faible. On est alors à la limite du critère de SHANNON qui interdit toute détection significative de dimension caractéristique de moins de 2 pixels. La fréquence spatiale associée est donnée par un rayon, pour le cercle dans la TF, grand comme la moitié d'un côté de son cadre (presque atteint). En-deçà de cette limite de 2 pixels, on s'expose à provoquer un repliement de spectre dont la présence brouille la lecture de la TF (bien souvent de façon subreptice pour l'utilisateur novice de la TF!). En conformité avec les remarques de la section précédente, c'est sans inquiétude que l'on note l'ampleur du rôle de la maille dans la TF à des échelles si petites dans l'espace direct : non seulement des lobes latéraux apparaissent dans la TF, et ce en dépit de l'échelle linéaire choisie pour la représenter, mais une plongée à l'oeil nu dans le motif lui-même laisse entrevoir une figure symptomatique d'un pavage hexagonal (cf rosace hexagonale ci-dessous).
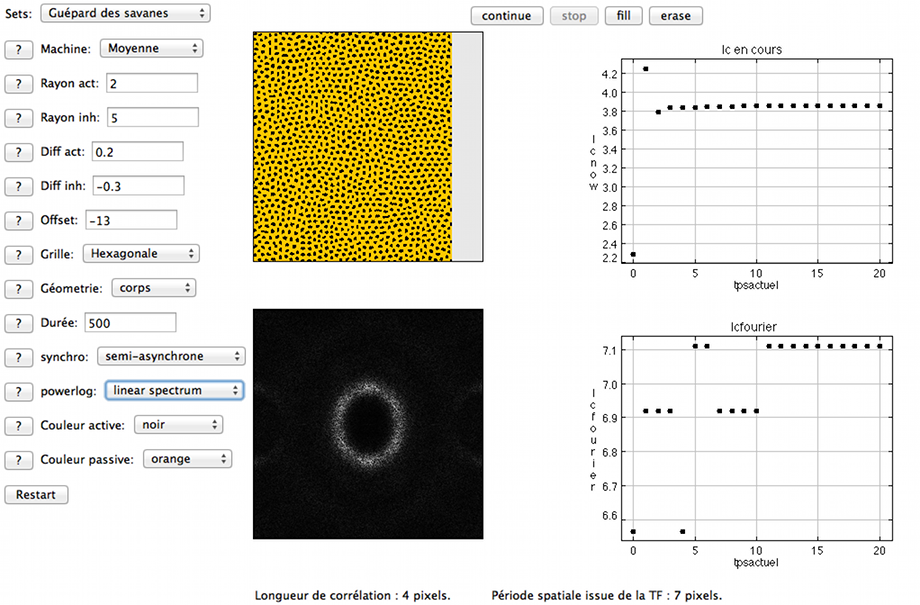
Configuration dérivée du Guépard des savanes, dans laquelle on réalise les déformations que peut introduire le maillage
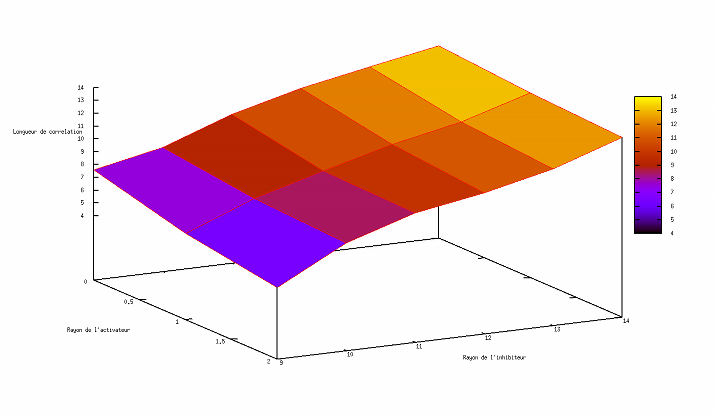
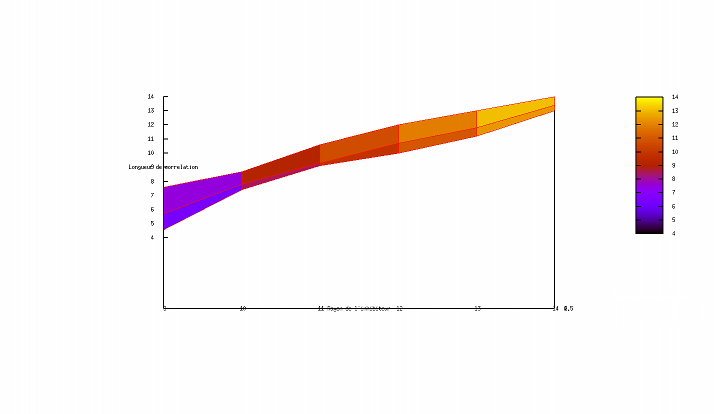
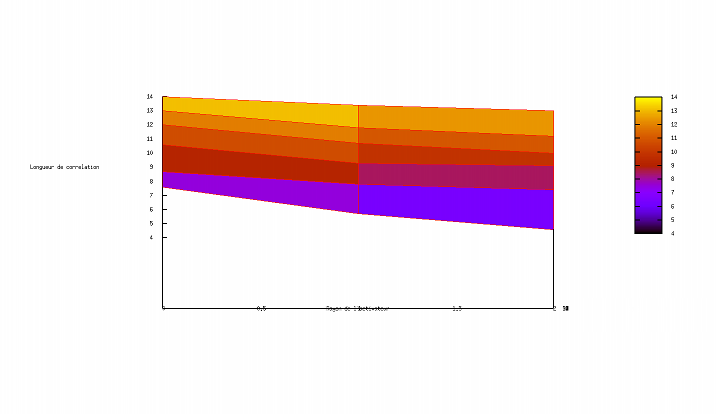
Exploration de l'espace des paramètres
Cette applet Java a avant tout un rôle pédagogique. Nous voulons croire qu'elle participe, à son modeste niveau, à illustrer les concepts parfois délicats qui sous-tendent les automates cellulaires et les équations de réaction-diffusion. Pourtant, un autre support numérique (eg un programme C++) serait l'occasion pour un lecteur curieux d'en savoir plus d'envisager une entreprise délicate pour une applet, une exploration de l'espace des paramètres. L'utilisateur se convaincra facilement de la sensibilité du motif aux paramètres ; il est facile de passer à côté de configurations pertinentes sans s'en apercevoir et difficile de prétendre avoir échantillonné de façon exhaustive sinon significative l'ensemble des configurations permises par ce formalisme (et les humbles capacités de calcul des machines).
Ne résistant pas à l'idée d'amorcer l'exploration d'un nouveau monde, nous nous sommes aventurés à esquisser une étude de corrélation entre l'évolution des rayons (activateur et inhibiteur) et la valeur de la longueur de corrélation associée (après stabilisation). Les résultats sont représentés ci-après :
Si les graphiques ci-dessus (réalisés avec une version plus ancienne de l'applet dans laquelle subsistait un formalisme plus archaïque) s'avèrent jamais avoir une vertu, ce sera bien celle d'avoir suscité la vocation exploratrice chez le chaland enthousiasmé. Une véritable exploration de l'espace des paramètres nécessite (du temps,) un talent et une expérience certaines pour anticiper les régions susceptibles d'être le lieu d'une découverte et appréhender les résultats pertinents.
Le marsupilami, un animal pas comme les autres
Un résultat célèbre obtenu par Alan TURING alors qu'il étudiait la morphégénèse, est illustré avec la configuration "Marsupilami corrigé". Avec son modèle, TURING parvint à expliquer pourquoi, alors que les mêmes mécanismes cellulaires locaux étaient à l'oeuvre sur l'ensemble du corps d'un animal comme le guépard, ce dernier présentait des taches circulaires sur le corps et des bandes orthogonales à la direction privilégiée sur la queue (cf figure ci-dessous). Dans la situation Marsupilami corrigé (comme toujours en géométrie "queue"), on a court-circuité les calculs de la fonction de corrélation et de la TF pour des raisons techniques. L'intérêt de cette configuration réside avant tout dans l'allure du motif en géométrie queue par rapport à celui obtenu, avec les mêmes paramètres, en géométrie corps. Bien que le mécanisme local d'activation des cellules reste le même, tout se passe comme si l'évolution des motifs dépendait de la géométrie globale du système! En effet, les mémes paramètres vont donner des bandes en géométrie queue et des taches en géométrie corps.
Remarquer la différence corps/queue
.jpg)
L'explication réside dans les conditions aux limites imposées au système. Ici, le terme queue est abusif en ce sens que, si la géométrie corps modélise bel et bien une sphère ( ~ corps réel de guépard ou de dalmatien), la géométrie queue correspond en fait à un tore (conditions au limites verticales mais aussi horizontales - le début et la fin de la queue sont reliés!). Néanmoins, la configuration "Marsupilami corrigé" a été réglée pour être sensible aux effets de bords haut/bas mais pas droite/gauche : on modélise bien la queue cylindrique d'un animal et on retrouve ainsi le célèbre résultat d'Alan TURING.

Motif dérivé de la configuration marsupilami (rayon de l'inhibiteur plus petit)
En diminuant le rayon de l'inhibiteur, on finit par perdre cette périodicité (les cellules ne se voient plus d'un bord à l'autre) et des taches réapparaissent sur la queue (cf queue ci-dessus). Ainsi, l'observation de la transition taches/bandes sur le pelage d'un animal ou dans un paysage renseigne sur le rayon de diffusion de l'entité qualifiée d'activatrice.
N.B. : l'amateur de bande dessiné n'aura pas manqué de remarquer que la queue du marsupilami original est tacheté, pas rayée, en dépit de taches similaires à celles du guépard sur le corps. Un bien étrange animal aux rayons fluctuants...
Animaux
Ci-dessous, on peut voir quelques exemples de pelage ou carapaces d'animaux dont la structure est proche de certaines simulations. Les images seront volontairement prises grandes (configuration
la plus puissante) pour le lecteur malchanceux dont la machine ne permettrait pas l'affichage de ces grandes configurations.
Guépard (Acinonyx Jubatus)


Les paramètres sont : R1=2, R2=10, J1=1, J2=-0.1, h=-18.
Guépard royal (Acinonyx jubatus rex)


Les paramètres sont : R1=5, R2=10, J1=1, J2=-0.1, h=0.
Hanneton Foulon (Polyphylla fullo)


Les paramètres sont : R1=3, R2=4, J1=6.5, J2=-3, h=0.
Cet animal, encore appelé hanneton des pins, prouve que les mammifères ne sont pas forcément les seuls concernés par le modèle de Turing.
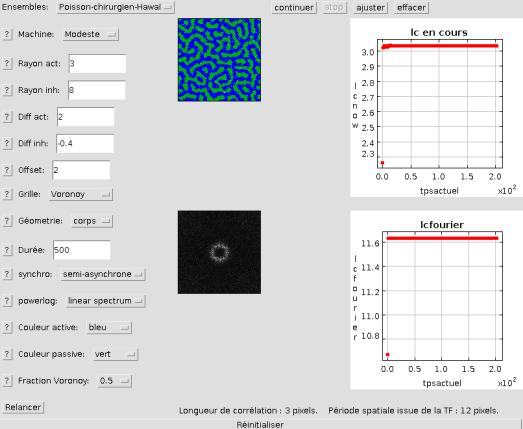
Poisson-chirurgien d'Hawaï
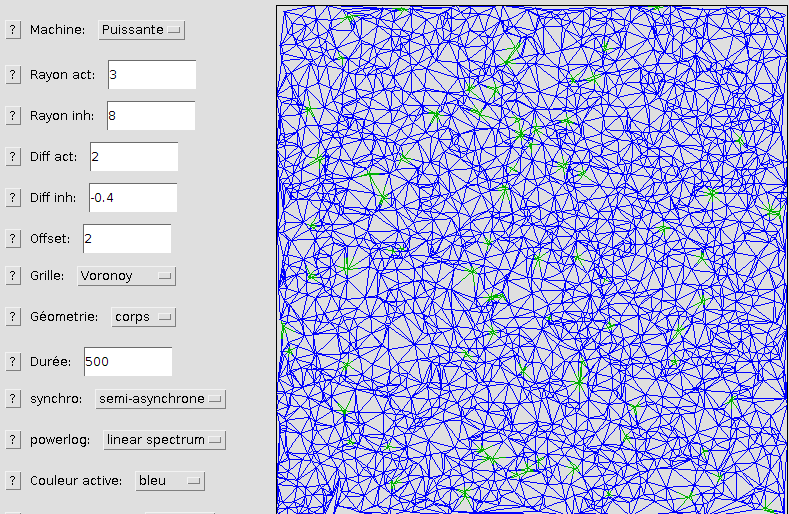
Ci-dessus, on peut observer l'apparence de l'ensemble ichtyen pour une densité faible de cellules de Voronoy. Cela permet d'observer l'agencement désordonné de la grille.
Ci-dessous, le même ensemble est généré avec la densité la plus élevée prévue. On voit que l'apparence est proche de celle des autres géométries (et du poisson)
avec une meilleure isotropie. Cela se vérifie sur la transformée de Fourier correspondante.
En observant la photographie du poisson, on remarque d'ailleurs une différence entre les motifs de sa tête, ses nageoires et son corps.
La simulation est plus proche d'une géométrie tête (avec ses conditions aux limites périodiques), d'où la plus forte ressemblance avec la tête de l'animal.
Dunes
Dunes du désert Namib en Namibie


Les paramètres sont : R1=2, R2=14, J1=0.5, J2=-0.1, h=0.
Certains géophysiciens ont appliqué le modèle de Turing avec succès à la formation de dunes (
voir ici).
Mode d'emploi de l'applet
Les Différents Types d'Affichage
L'utilisateur a le choix entre quatre types d'affichage principaux : "Motif et corrélation", "Motif et FFT", "Motif seul" et "Section de la TF".
Pour le premier, l'applet Java affiche l'image de la grille où se développe la structure de Turing avec trois courbes. En haut à droite est tracée la fonction de corrélation de l'image définie précédemment. En bas à gauche est reportée la valeur de la taille caractéristique déterminée à partir de la fonction de corrélation en fonction du numéro de l'itération (du temps). En bas à droite est tracée la somme de toutes les cellules de la grille : toujours avec la convention
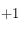
pour une cellule activée et
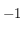
pour une cellule désactivée. Ce graphe est intéressant pour se donner une idée de l'instant où le système converge vers un état stable où plus aucune cellule ne change d'état : lorsque la valeur de la somme devient constante.
Cela étant dit, la constance de la valeur de la somme au fil du temps n'implique pas nécessairement que plus aucune cellule ne change d'état (permutation entre le même nombre de cellules activées et désactivées). Cependant, dans la grande majorité des cas, cette affirmation est justifiée. Pour s'en convaincre, il suffit de regarder l'évolution de la valeur calculée de la taille caractéristique qui devient elle aussi constante lorsque le système converge vers un état stable.
Concernant le deuxième affichage "Motif et FFT", il produit comme le précédent l'image de la grille où se développe la structure de Turing mais aussi la transformée de Fourier de cette image (panneau en bas à gauche). L'idée est alors de calculer la taille caractéristique du système grâce à deux méthodes indépendantes. Ces valeurs sont reportées dans les panneaux à droite de l'affichage : en haut se trouve la taille caractéristique calculée à partir de la fonction de corrélation (identique au premier affichage), en bas est reportée la taille caractéristique calculée à partir de la transformée de Fourier.
La concordance entre les valeurs retournées de la taille caractéristique par ces deux méthodes assure une certaine crédibilité quant au chiffre avancé.
Le troisième et le quatrième affichage permettent à l'utilisateur de se focaliser sur une partie précise des deux affichages principaux. "Motif seul" n'affiche que la grille où les taches évoluent alors que "Section de la TF" affiche la moyenne des points de la transformée de Fourier situés dans des anneaux centrés sur l'origine de la TF (voir la Section sur la Transformée de Fourier).
Les Coefficients
L'utilisateur a aussi la possibilité de modifier plusieurs coefficients et laisser évoluer le système pour faire apparaître des structures. Certaines de ces structures correspondent au pelage, à la carapace... de certains animaux. Les ensembles de coefficients responsables de l'émergence de ces structures ont été répertoriés et rassemblés dans l'onglet "ensemble". L'utilisateur peut alors tester les différentes combinaisons proposées et éventuellement les comparer à des images représentant les animaux correspondants.
Les coefficients peuvent être modifiés séparément laissant ainsi libre cours à l'utilisateur de trouver de nouvelles combinaisons. Ces coefficients sont :
-
"Machine" : suivant la puissance de l'ordinateur à partir duquel est lancé la simulation, ce paramètre permet de choisir la taille de la grille contenant les cellules que l'on souhaite faire évoluer. Il y a trois choix possibles : Modeste (128*128 pixels), Moyenne (256*256 pixels) et Puissante (512*512 pixels). Il est conseillé à l'utilisateur de ne pas tenter l'affichage "Machine Puissante" si il ne dispose pas d'un relativement bon ordinateur.
Choix "Machine"
-
"Rayon act" : définit la valeur en pixels du rayon autour duquel la cellule activée envoie des composés chimiques responsable de l'activation des cellules. Ce rayon est inférieur au rayon inhibiteur.
-
"Rayon inh" : définit la valeur en pixels du rayon autour duquel la cellule activée envoie des composés chimiques responsable de l'inhibition, la désactivation des cellules.
Choix des Rayons d'Actions
-
"Diff act" : définit la "force" des composés chimiques responsables de l'activation des cellules.
-
"Diff inh" : définit la "force" des composés chimiques responsables de l'inhibition des cellules.
Choix des Coefficients de Diffusion
-
"Offset" : définit la valeur (somme de "Diff act" et de "Diff inh" pour une cellule) à partir de laquelle une cellule change d'état.
Choix de l'Offset

-
"Grille" : est un paramètre permettant de choisir la nature de la grille à partir de laquelle les cellules sont formées. Il y a trois choix possibles : Simple (un maillage carré standard), Hexagonale et Voronoi (une structure plus complexe définie dans la page consacrée aux maillages, voir ci-dessus).
Choix de la Grille
-
"Géométrie" : est un autre paramètre offrant la possibilité de choisir la forme géométrique de la grille. On a vu que les structures de Turing sont dépendantes de la géométrie de la surface sur laquelle elles se forment pour un même ensemble de paramètres.
Choix de la Géométrie de Grille
-
"Durée" : ce paramètre n'a de sens que dans le cas du choix d'évolution "asynchrone" et définit le nombre de cellules mises à jour avant l'affichage. Il influence directement le temps de calcul de l'applet. Il est conseillé à l'utilisateur, s'il veut voir l'évolution de la grille à "échelle humaine", de ne pas choisir de durée trop faible. Typiquement, la valeur optimale est 500 mais il est tout à fait possible de diminuer le temps d'évolution en choisissant une durée de 1000 voire 10000.
Choix du Nombre d'Itérations avant Affichage
-
"Synchro" : permet de choisir le mode de mise à jour de la grille : il y a trois choix possibles qui ont été définis précédemment : asynchrone, synchrone et semi-asynchrone.
Choix du type d'évolution de la Grille
-
"Powerlog" : permet de choisir le type d'affichage de la Transformée de Fourier : soit linéaire soit logarithmique.
Choix de l'Affichage de la Transformée de Fourier
-
"Couleur active" : élément définissant la couleur des cellules activées.
-
"Couleur passive" : élément définissant la couleur des cellules désactivées.
Choix de la Couleur d'affichage
Exemple d'utilisation
Pour aider le lecteur à tirer tout le parti de cette appliquette, voici un petit exemple commenté.
On peut tout simplement choisir un ensemble de paramètres prédéfini pour ressembler à un animal. Prenons pour les besoins de cet exemple, le hanneton foulon, un insecte au corps tacheté.
Choix d'un ensemble
On voit alors apparaître dans les champs les valeurs prédéfinies correspondant à l'ensemble. Elles peuvent encore être modifiées si nécessaire. Ici nous ne le ferons pas.
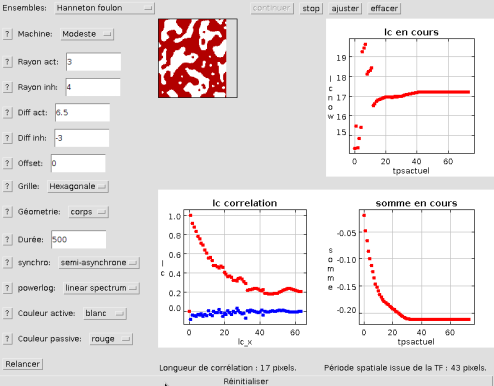
C'est le moment de cliquer sur "Motif et corrélation" pour voir évoluer en direct les cellules. Avant le premier affichage peut s'écouler un peu de temps.
Cela correspond à l'initialisation de la grille, et de ses sous-objets numériques associés (nous n'entrerons pas plus avant dans les détails). Normalement après quelques itérations vous pourrez voir ceci à l'écran:
Cette configuration ressemble beaucoup à cette photographie de l'animal:
Hanneton Foulon

Outre les taches évoluant en direct, on peut voir en dessous de celles-ci (en bas à gauche) la courbe de la fonction de corrélation calculée sur la figure.
Tiré de cette courbe de corrélation, en haut à droite est placé un graphe présentant la distance caractéristique correspondante au fil de temps. En bas à droite, utile pour vérifier si l'automate est stabilisé ou non, on peut observer la courbe de la somme totale sur la grille en fonction de l'itération (pondérée par le nombre de cellules).
On peut ensuite, pour voir un autre aspect de l'analyse, réinitialiser l'appliquette (bouton du bas
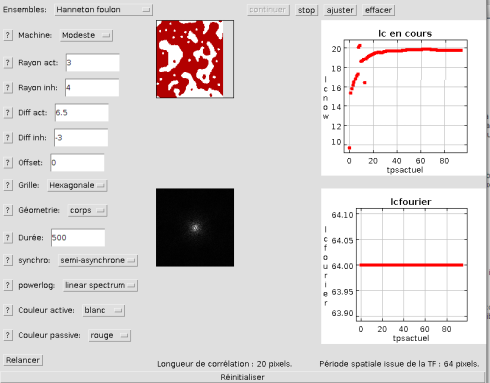
). Nous allons maintenant choisir le deuxième bouton possible ("Motif et FFT").
Celui-ci présente en parallèle à gauche l'image et sa transformée de Fourier à deux dimensions. A droite on peut voir de haut en bas la longueur caractéristique calculée à partir de la corrélation, et celle calculée à partir de la transformée de Fourier.
La longueur caractéristique calculée par corrélation est de 20 pixels et celle calculée par transformée de Fourier de 64 pixels pour cette réalisation.
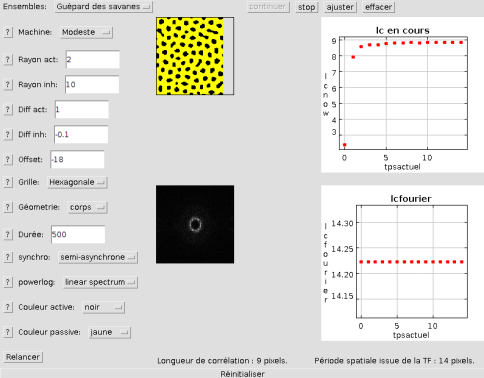
Pour le guépard, on trouve, comme on le voit ci-dessous, une meilleure concordance car les deux méthodes ne sont pas calibrées pour détecter la même chose.
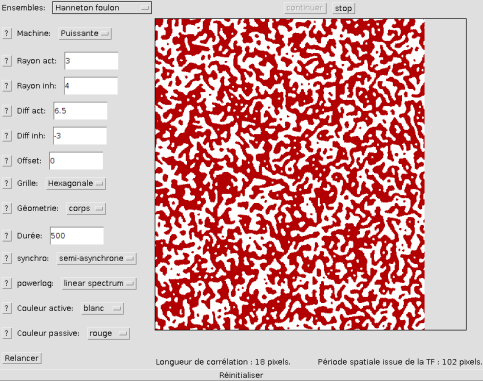
Si l'on veut observer une zone, la plus grande possible, de structures simulées, il convient de choisir le bouton "Motif seul" (et de choisir une machine plus puissante si possible).
Ainsi les taches se montreront dans toute leur splendeur sur tout l'espace disponible:
N'hésitez pas à essayer tous les animaux prédéfinis par vous-même! Et à expérimenter avec différentes valeurs numériques. Vous pourrez même choisir la couleur qui leur convient le mieux!
Le modèle pourrait encore être perfectionné pour expliquer les complexités de pelages comme celui de la magnifique panthère nébuleuse... Notons que celle-ci
présente également une dichotomie entre les parties de son corps à géométrie différente... Notons également qu'elle fait partie de la liste des
espèces menacées.
Panthère nébuleuse

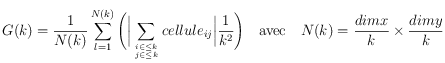

{kind=link}